Gut Microbiota Composition Can Shape Mouse VOC Breath Profiles
The gut microbiota is associated with multiple roles in health and disease and is responsible for a number of VOCs that are detectable in breath.
| Publication information: Hernandez-Leyva AJ, Berna AZ, Liu Y, Rosen AL, Lint MA, Whiteside SA, Jaeger N, McDonough RT, Joardar N, Santiago-Borges J, Tomera CP. The breath volatilome is shaped by the gut microbiota. medRxiv. 2024 Aug 8.
Research area: Gut Microbiome Sample medium: Breath, fecal sampling Analysis approach: GC-qToFMS Summary:
|
Introduction
Analysis of the VOC content of exhaled breath as biomarkers has marked advantages for clinical research and practice. This includes the inherent non-invasive collection of breath as a sampling medium, the direct and rapid link between VOC production in the body from metabolic pathways and their detection in the breath, and the inexhaustible nature of breath. We have previously shown that a significant number of VOCs detectable in the breath are thought to originate from the gut microbiome.
Many microbial metabolites already have strong links to specific species, pathways, and processes (1–6), many of which happen to be volatile. Studies have demonstrated a strong, significant, relationship between the composition of the intestinal microbiota and the VOCs that are detectable in exhaled breath (7). For example, dietary fibers, which human metabolism cannot process, are fermented by microbes in the intestines, producing VOCs such as short-chain fatty acids (SCFAs).
These VOCs are thought to have roles in several signaling contexts including the central nervous system, immunity, and inflammation (8–10). To progress the development of breath VOC biomarkers of gut microbes and their involvement in human health, identifying to what degree VOC changes in the breath reflect changes in the gut microbiota is important.
Previous research has demonstrated the connection between exhaled breath VOCs and intestinal microbiota in subjects with Crohn’s disease (CD) (7). In this study, breath samples were collected and analyzed in the same subjects during both active symptom flares and during remission. The SCFAs acetate and propionate correlated with Bifidobacteria and several other microbes in the Firmicutes phylum in both disease states. Also, the microbial strains and the relative abundances of SCFAs decreased in the active disease state.
To further investigate how the gut microbiome shapes the VOCs detectable in the breath, Hernandez-Leyva et al., compared the breath VOC profile of 27 healthy children to the microbiota species detectable in paired stool samples (11). Next, to investigate potential causal relationships between the two, a gnotobiotic mouse model was developed to enable robust control of the gut microbiome. Through this method, the VOCs emitted from cultured gut microbes grown under anaerobic conditions were able to be compared to the VOCs detectable in the breath of mice colonized with the same bacterial strains.
Breath Collection Methods
A total of 27 healthy children between 6 – 12 years of age in the Microbiome-Asthma Connection in Kids (MACK) study with matched stool and breath samples were utilized as part of this study. Breath was sampled through subjects exhaling into a disposable cardboard mouthpiece connected to a chamber, which was then pulled through thermal desorption tubes for GC-MS analysis. Mice were transferred into sterilized isolators and maintained on the same autoclaved chow and water as germ-free mice for one week before the start of the experiment.
Monocolonized gnotobiotic mice were kept separately in a specialized facility. Mouse breath was collected using a ventilator directly from the respiratory system via tracheostomy, which bypasses the oral microbiome. To collect a clean background equipment sample, the sample path of the ventilator was blocked prior to mouse breath sampling and 3 volumes of 1.1L of air were collected. After five minutes of venting the output line with mouse breath, the output port of the ventilator was then connected to a bag for breath collection. The water vapor in the bags was allowed to condense for 15 minutes, and then the air was drawn through thermal desorption tubes for later analysis via TD-GC-MS.
Results
VOCs in the breath correlate with gut microbiome composition in healthy children.
The gut microbiome was characterized in fecal samples through whole metagenomic sequencing, allowing the identification of different phyla. The composition of the gut microbiome was consistent with what has been established as normal previously, with a dominance of Bacillota, Bacteriodota, and Actinomycetota, and the presence of Pseudomonadota, Fusobacteriota, and Verrucomicrobiota. Next, breath samples were analyzed using gas chromatography and time-of-flight mass spectrometry (GC-qToFMS).
A total of 2734 features were detected, and statistical analysis was used to investigate whether the differences in VOC content between children correlated with the differences in gut microbiota composition. The gut metagenome (P = 0.023), uniref90 gene clusters (P = 0.011), and functional pathways (P = 0.03) were significantly correlated with breath VOC composition.
From this, 51 molecular features were chemically identified as VOCs, including acetic acid (a short-chain fatty acid), camphene, 3-carene, beta-pinene, limonene, and p-cymene. Statistical analysis was used to assess whether specific bacterial taxa correlated with these named VOCs. A substantial abundance variance (up to 40%) of these selected breath VOCs was explained by composition changes in the gut microbiota. Further analysis revealed particularly strong associations between several bacteria and common breath VOCs.
VOCs have mechanistic associations with gut microbiome species in gnotobiotic mice models
To make more mechanistic assessments of the gut microbial origins of breath VOCs, the breath and fecal samples of mice with distinct gut microbiota compositions due to being sourced from different vendors (12) were collected. To also test whether breath VOCs could be transferred by gut microbiota transplant, stool samples from conventional mouse groups were homogenized and used to colonize the gastrointestinal tract of sex-matched germ-free mice (Figure 1).
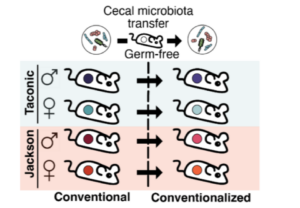
Figure 1 – Overview of a gnotobiotic animal model for testing the contribution of the gut microbiota to the breath volatilome (taken from Hernandez-Leyva et al., 2024).
Mouse breath was collected using a ventilator through a cannula surgically implanted into the trachea to maximize the collection of VOCs from the alveolar air. Similar to the results found in human breath, mouse breath VOC composition correlated with the gut microbiota, including taxonomic profile (P = 0.0047), and uniref90 cluster compositions (P = 0.006). The same curated list of 51 VOCs identified in human breath was used to investigate their microbial associations in mice.
A significant number of breath VOCs varied between different experimental groups of mice with different gut microbiota compositions including benzene and ethyl acetate, indicating that the microbiota with or without transplant impacted the abundance of VOCs detectable in breath. The top five breath VOCs best explained by gut microbiota composition (benzene, levomenthol, 4-methyl-2-pentanone, linalool, and 2-ethylhexan-1-ol) all had greater than a 40% mean variance explained, interestingly, several breath VOCs shared a relatively high variance explained by the gut microbiota in both humans and mice.
To make mechanistic conclusions about specific bacterial species in the gut and breath VOCs, the VOC profile between bacterial culture headspace, and mice monocolonized with the same bacterial species were compared (Figure 2).
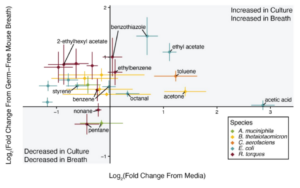
Figure 2 – Comparing the volatilome of anaerobic culture headspace to the exhaled breath of monocolonized mice identifies potential gut microbiota-derived breath VOCs (taken from Hernandez-Leyva et al., 2024).
E. coli showed an association in culture and in the breath of mice with benzothiazole, ethyl acetate, and octanal, B. thetaiotaomicron was associated with acetone, and C. aerofaciens was associated with toluene. Pentane was reduced in both the bacterial culture headspace, and in the breath of mice colonized with R. torques. Together, these results suggest that not only do distinct microbial communities in the gut influence the VOC composition of breath, but that individual VOCs can be associated with specific bacterial species.
Discussion
The results of this study validate the findings of previous studies that indicate that gut microbiome species composition, as well as functional pathways are correlated to breath VOC composition. Previous studies investigating the association between breath VOCs and gut microbiota have mostly been conducted in human subjects, examining changes to the diet or distinguishing between health and disease. However, these foundational studies offer limited control over many of the factors that affect the breath volatilome composition, such as diet and environmental exposures (60).
This provides new evidence linking variation in the gut microbiota to changes in the breath volatilome of the host, in both mouse and humans. Moreover, this study extended these findings by indicating that specific bacterial species can be associated with individual breath VOCs through using gnotobiotic mice and paired bacterial cultures. This also demonstrated the power of using mouse models in breath VOC biomarker research. Aligned with previous literature, many of the most microbially-associated breath VOCs were terpenes.
Terpenes are a diverse chemical class that are able to be synthesized by plants and microbes, but not mammalian metabolism. The terpenes identified in this study included camphene, 3-carene, beta-pinene, limonene, and p-cymene. The microbial-produced VOCs, including terpenes, are suspected to influence human physiology through multiple mechanisms. We have previously associated levels of limonene in the breath with liver function, and have designed a dynamic breath test that can assay liver function for the diagnosis of liver cirrhosis (13,14).
We have also recently conducted a literature review of the VOCs associated with malaria, and many terpenes were also shown to be associated, including alpha-pinene, limonene, beta-pinene, 3-carene, terpinolene, and m-cymene. Overall, this study specified the connections between certain gut microbes and the VOCs they produce that are detectable in breath. This provides novel insights into some of the long-suspected VOC origins in exhaled breath.
How can Owlstone Medical help with your gut microbiome research?
We have many tools and services that can incorporate breath analysis into microbiome research, including our handheld hydrogen and methane device, and Breath Biopsy Collection Station. Why not download our microbiome content pack to get you started on discovering what microbiome-associated VOCs have been reliably detected in the breath, and access all of our tailored microbiome content in one place?
The Collection Station is part of Breath Biopsy OMNI® our end-to-end optimized service for consistent breath analysis. Another aspect of OMNI is access to our Breath Biopsy VOC Atlas®. Currently more than 200 rigorously identified VOCs are registered within the ATLAS, including gut microbiome-associated VOCs like sulfurous compounds, short-chain fatty acids (SCFAs) like acetate, butyrate, and propionate, and aromatic amino acid breakdown products. Join the waitlist to again access as soon as possible.
Our team is always willing to discuss with you to discover whether breath analysis could support your research aims. Please do not hesitate to contact us via our website to get started.
References
- Le Gall G, Noor SO, Ridgway K, Scovell L, Jamieson C, Johnson IT, et al. Metabolomics of Fecal Extracts Detects Altered Metabolic Activity of Gut Microbiota in Ulcerative Colitis and Irritable Bowel Syndrome. J Proteome Res. 2011 Sep 2;10(9):4208–18. doi: 10.1021/pr2003598
- Kim S, Yin X, Prodhan MAI, Zhang X, Zhong Z, Kato I. Global Plasma Profiling for Colorectal Cancer-Associated Volatile Organic Compounds: a Proof-of-Principle Study. J Chromatogr Sci. 2019 May;57(5):385–96. doi: 10.1093/chromsci/bmz011
- Huang H jun, Zhang A ye, Cao H cui, Lu H feng, Wang B hong, Xie Q, et al. Metabolomic analyses of faeces reveals malabsorption in cirrhotic patients. Dig Liver Dis. 2013 Aug 1;45(8):677–82. doi: 10.1016/j.dld.2013.01.001
- Xie G, Wang X, Huang F, Zhao A, Chen W, Yan J, et al. Dysregulated hepatic bile acids collaboratively promote liver carcinogenesis. Int J Cancer. 2016;139(8):1764–75. doi: 10.1002/ijc.30219
- Chen MX, Wang SY, Kuo CH, Tsai IL. Metabolome analysis for investigating host-gut microbiota interactions. J Formos Med Assoc. 2019 Mar 1;118:S10–22. doi: 10.1016/j.jfma.2018.09.007
- Obrenovich ME, Tima M, Polinkovsky A, Zhang R, Emancipator SN, Donskey CJ. Targeted Metabolomics Analysis Identifies Intestinal Microbiota-Derived Urinary Biomarkers of Colonization Resistance in Antibiotic-Treated Mice. Antimicrob Agents Chemother. 2017 Jul 25;61(8):e00477-17. doi: 10.1128/AAC.00477-17
- Smolinska A, Tedjo DI, Blanchet L, Bodelier A, Pierik MJ, Masclee AAM, et al. Volatile metabolites in breath strongly correlate with gut microbiome in CD patients. Anal Chim Acta. 2018 Sep 26;1025:1–11. doi: 10.1016/j.aca.2018.03.046
- Frost G, Sleeth ML, Sahuri-Arisoylu M, Lizarbe B, Cerdan S, Brody L, et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat Commun. 2014 Apr 29;5:3611. doi: 10.1038/ncomms4611
- Parada Venegas D, De la Fuente MK, Landskron G, González MJ, Quera R, Dijkstra G, et al. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front Immunol. doi: 10.3389/fimmu.2019;277.
- Cronin P, Joyce SA, O’Toole PW, O’Connor EM. Dietary Fibre Modulates the Gut Microbiota. Nutrients. 2021 May 13;13(5):1655. doi: 10.3390/nu13051655
- Hernandez-Leyva AJ, Berna AZ, Liu Y, Rosen AL, Lint MA, Whiteside SA, et al. The breath volatilome is shaped by the gut microbiota. MedRxiv Prepr Serv Health Sci. 2024 Aug 8;2024.08.02.24311413. doi: 10.1101/2024.08.02.24311413
- Rasmussen TS, de Vries L, Kot W, Hansen LH, Castro-Mejía JL, Vogensen FK, et al. Mouse Vendor Influence on the Bacterial and Viral Gut Composition Exceeds the Effect of Diet. Viruses. 2019 May 13;11(5):435. doi: 10.3390/v11050435
- Ferrandino G, De Palo G, Murgia A, Birch O, Tawfike A, Smith R, et al. Breath Biopsy® to Identify Exhaled Volatile Organic Compounds Biomarkers for Liver Cirrhosis Detection. J Clin Transl Hepatol. 2023 Jun 28;11(3):638–48. doi: 10.14218/JCTH.2022.00309
- Ferrandino G, Ricciardi F, Murgia A, Banda I, Manhota M, Ahmed Y, et al. Exogenous Volatile Organic Compound (EVOC®) Breath Testing Maximizes Classification Performance for Subjects with Cirrhosis and Reveals Signs of Portal Hypertension. Biomedicines. 2023 Nov;11(11):2957. doi: 10.3390/biomedicines11112957
Catch up on the presentations from the Breath Biopsy Conference 2024
